
Оглавление:
Оформленное клеточное ядро это




Почитать еще
Эукариоты — это истинно-ядерные организмы, клетки которых имеют оформленное клеточное ядро, отделенное от цитоплазмы ядерной оболочкой. Для огромного большинства их характерно деление ядра с образованием настоящих хромосом и половой процесс, при котором образуются ядра с уменьшенным вдвое (редуцированным) числом хромосом.
Хромосомы эукариот связаны с особыми, гистоновыми белками. Содержание ДНК в них в десятки раз больше, чем это необходимо для кодирования белков.
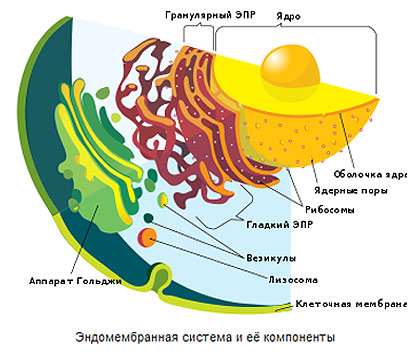
Эндомембранная система и её компоненты
Эукариотические клетки часто имеют настоящие жгутики, сложенные из волокон — фибрилл, расположенных по схеме 9+2 — девять по окружности и две в центре. Важная особенность эукариот — наличие в их цитоплазме клеточных органоидов, имеющих свой небольшой геном (совокупность генов) и размножающихся делением. Это митохондрии, а у высших растений — хлоропласт. Возможно, они произошли из клеточных симбионтов: первые — от каких-то дышащих кислородом (аэробных) бактерий, а вторые — из цианобактерий. Все клетки, имеющие митохондрии, дышат кислородом, лишь немногие эукариоты потеряли способность к аэробному обмену.
В надцарство эукариот входят разнообразные простейшие (жгутиковые, инфузории, споровики), грибы, животные и растения. Эукариотам противопоставляют более примитивную группу — прокариот.
Источник: Энциклопедический словарь юного биолога. Составитель Аспиз М. Е. Издательство «Педагогика», Москва, 1985
zooschool.ru
ЯДРО (клеточное ядро), в биологии — обязательная часть клетки у многих одноклеточных и всех многоклеточных организмов. Размеры от 1 мкм (у некоторых простейших) до 1 мм (в яйцах некоторых рыб и земноводных). Все организмы нашей биосферы как одноклеточные, так и многоклеточные, подразделяются на эукариот— их клетки содержат ядро, и прокариот, клетки которых не имеют морфологически оформленного ядра. Термин «ядро» (лат. nucleus ) впервые применил Р. Броун в 1833 году, когда описывал шарообразные структуры, наблюдаемые им в клетках растений.
Внутреннее пространство клеточного ядра отделено от цитоплазмы ядерной оболочкой, состоящей из двух мембран. Мембраны оболочки ядра сходны по строению с другими мембранными компонентами клетки и построены по тому же принципу: это тонкие липопротеидные пленки, состоящие из двойного слоя липидных молекул, в который встроены молекулы белков. Пространство между внутренней и внешней ядерными мембранами называется перинуклеарным. На поверхности внешней ядерной мембраны обычно располагается большое количество рибосом, и иногда удается наблюдать непосредственный переход этой мембраны в систему каналов гранулярной эндоплазматической сети клетки. Внутренняя ядерная мембрана связана с тонким волокнистым белковым слоем — ядерной ламиной, состоящей из белков ламинов. Густая сеть фибрилл ядерной ламины способна обеспечить целостность ядра, даже после растворения липидных мембран оболочки ядра в эксперименте. С внутренней стороны к ламине крепятся петли хроматина, заполняющего ядро.
Ядерная оболочка имеет отверстия диаметром около 90 нм, образующиеся засчет слияния внешней и внутренней ядерных мембран. Такие отверстия в оболочке ядра окружены сложными белковыми структурами, получившими название комплекса ядерной поры. Восемь белковых субъединиц, входящих в состав ядерной поры, располагаются вокруг перфорации ядерной оболочки в виде колец, диаметром около120 нм, наблюдаемых в электронный микроскоп с обеих сторон ядерной оболочки. Белковые субъединицы комплекса поры имеют выросты, направленные к центру поры, где иногда видна «центральная гранула» диаметром 10-40 нм. Размер ядерных пор и их структура стандартны для всех клеток эукариот. Число ядерных пор зависит от метаболической активности клеток: чем выше уровень синтетических процессов в клетке, тем больше пор на единицу площади поверхности клеточного ядра. В процессе ядерно-цитоплазматического транспорта ядерные поры функционируют как некое молекулярное сито, пропуская ионы и мелкие молекулы (сахара, нуклеотиды, АТФ и др.) пассивно, по градиенту концентрации, и осуществляя активный избирательный транспорт крупных молекул белков и рибонуклеопротеидов, то есть комплексов рибонуклеиновых кислот (РНК) с белками. Так, например, белки, транспортируемые в ядро из цитоплазмы, где они синтезируются, должны иметь определенные последовательности примерно из 50 аминокислот, (т. наз. NLS последовательности), «узнаваемые» комплексом ядерной поры. В этом случае комплекс ядерной поры, затрачивая энергию в виде АТФ, активно транслоцирует белок из цитоплазмы в ядро.
Клеточное ядро является вместилищем практически всей генетической информации клетки, поэтому основное содержимое клеточного ядра — это хроматин: комплекс дезоксирибонуклеиновой кислоты (ДНК) и различных белков. В ядре и, особенно, в митотических хромосомах, ДНК хроматина многократно свернута, упакована особым образом для достижения высокой степени компактизации. Ведь все длинные нити ДНК, общая длина которых составляет, например, у человека около 164 см, необходимо уложить в клеточное ядро, диаметр которого всего несколько микрометров. Эта задача решается последовательной упаковкой ДНК в хроматине с помощью специальных белков. Основная масса белков хроматина — это белки гистоны, входящие в состав глобулярных субъединиц хроматина, называемых нуклеосомами. Всего существует 5 видов белков гистонов. Нуклеосома представляет собой цилиндрическую частицу, состоящую из 8 молекул гистонов, диаметром около 10 нм, на которую «намотано» чуть менее двух витков нити молекулы ДНК. В электронном микроскопе такой искусственно деконденсированный хроматин выглядит как «бусины на нитке». В живом ядре клетки нуклеосомы плотно объединены между собой с помощью еще одного линкерного гистонового белка, образуя так называемую элементарную хроматиновую фибриллу, диаметром 30 нм. Другие белки, негистоновой природы, входящие в состав хроматина обеспечивают дальнейшую компактизацию, т. е. укладку, фибрилл хроматина, которая достигает своих максимальнах значений при делении клетки в митотических или мейотических хромосомах. В ядре клетки хроматин присутствует как в виде плотного конденсированного хроматина, в котором 30 нм элементарные фибриллы упакованы плотно, так и в виде гомогенного диффузного хроматина. Количественное соотношение этих двух видов хроматина зависит от характера метаболической активности клетки, степени ее дифференцированности. Так, например, ядра эритроцитов птиц, в которых не происходит активных процессов репликации и транскрипции, содержат практически только плотный конденсированный хроматин. Некоторая часть хроматина сохраняет свое компактное, конденсированное состояние в течение всего клеточного цикла — такой хроматин называется гетерохроматином и отличается от эухроматина рядом свойств.
Репликация и транскрипция
Клетки эукариот содержат обычно несколько хромосом (от двух до нескольких сотен), которые теряют в ядре (в интерфазе, т. е. между митотическоми делениями) клетки свою компактную форму, разрыхляются и заполняют объем ядра в виде хроматина. Несмотря на деконденсированное состояние, каждая хромосома занимает в ядре строго определенное положение и связана с ядерной оболочкой посредством ламины. Строго закреплены на внутренней поверхности оболочки ядра такие структуры хромосом, как центромеры и теломеры. На определенной стадии жизненного цикла клетки, в синтетическом периоде, происходит репликация, т. е. удвоение всей ДНК ядра, и хроматина становится в два раза больше. Белки, необходимые для этого процесса, поступают, конечно, из цитоплазмы через ядерные поры. Таким образом, клетка готовится к предстоящему клеточному делению — митозу, когда общее количество ДНК в ядре вернется к первоначальному уровню.
Реализация генетической информации, заключенной в ДНК в виде генов, начинается с транскрипции, т. е. с синтеза информационных РНК (и-РНК) — точных копий генов, по которым затем будут строиться в цитоплазме на рибосомах белки. Этот процесс проходит в различных точках в обьеме ядра, морфологически ничем не отличающихся от окружающего хроматина. Чаще всего удается наблюдать транскрипцию диффузного, т.е. деконденсированного хроматина.
Кроме хроматина, составляющего хромосомы, в ядрах эукариот обычно содержится одно или несколько ядрышек. Это плотные структуры, не имеющие собственной оболочки и представляющие собой скопления молекул другого типа РНК — рибосомной РНК (р-РНК) в комплексе с белками. Такие комплексы называют рибонуклеопротеидами (РНП). Ядрышки имеют стандартную морфологию и образуются в ядре после деления клетки вокруг постояннодействующих точек активного синтеза рибосомной РНК. Гены рибосомной РНК, в отличие от большинства других генов, кодирующих белки, содержатся в геноме в виде многочисленных копий. Эти копии, расположенные в молекуле ДНК тандемно, т. е. друг за другом, располагаются в определенных районах нескольких хромосом генома. Такие районы хромосом называют ядрышковыми организаторами. Морфологически в ядрышке с помощью электронного микроскопа можно выделить следующие 3 зоны: гомогенные компактные фибриллярные центры, содержащие ДНК ядрышковых организаторов; плотный фибриллярный компонент вокруг них, где идет транскрипция генов рибосомной РНК и массивный гранулярный компонент ядрышка, состоящий из частиц РНП — будущих рибосом. Эти гранулы РНП, образующиеся в ядрышке, транспортируются в цитоплазму и образуют рибосомы, осуществляющие синтез всех белков клетки. Третий основной тип клеточных РНК — мелкие транспортные РНК — транскрибируются в различных участках ядра и выходят в цитоплазму через ядерные поры. Там они, как известно, обеспечивают транспортировку аминокислот к рибосомам в процессе синтеза белков.
Ядерный белковый матрикс
Для осуществления процессов репликации, транскрипции, а также поддержания определенного положения хромосом в обьеме ядра существуют каркасные белковые структуры, называемае ядерным белковым матриксом. Такой матрикс состоит, по крайней мере из трех морфологических компонентов: периферического фиброзного слоя- ламины; внутреннего, или интерхроматинового матрикса ядра и матрикса ядрышка. Наблюдения показывают, что компоненты ядерного матрикса — это не жесткие застывшие структуры, они динамичны и могут сильно видоизменяться в зависимости от функциональных особенностей ядер. Показано, что белковый матрикс имеет множество точек прочного связывания с ДНК ядра, которая, в свою очередь, имеет специальные последовательности нуклеотидов, необходимые для этого.
gimn1567.ru
Клеточное ядро


Ядро (лат. nucleus ) — это один из структурных компонентов эукариотической клетки, содержащий генетическую информацию (молекулы ДНК), осуществляющий основные функции: хранение, передача и реализация наследственной информации с обеспечением синтеза белка. Ядро состоит из хромати́на, я́дрышка, кариопла́змы (или нуклеоплазмы) и ядерной оболочки. В клеточном ядре происходит репликация (или редуплика́ция) — удвоение молекул ДНК, а также транскрипция — синтез молекул РНК на молекуле ДНК.
Содержание
Тонкая структура клеточного ядра

Огромная длина молекул ДНК эукариот предопределила появление специальных механизмов хранения, репликации и реализации генетического материала. Хроматином называют молекулы хромосомной ДНК в комплексе со специфическими белками, необходимыми для осуществления этих процессов. Основную массу составляют «белки хранения», так называемые гистоны. Из этих белков построены нуклеосомы — структуры, на которые намотаны нити молекул ДНК. Нуклеосомы располагаются довольно регулярно, так что образующаяся структура напоминает бусы. Нуклеосома состоит из белков четырех типов: H2A, H2B, H3 и H4. В одну нуклеосому входят по два белка каждого типа — всего восемь белков. Гистон H1, более крупный чем другие гистоны, связывается с ДНК в месте ее входа на нуклеосому. Нуклеосома вместе с H1 называется хроматосомой.
Нить ДНК с нуклеосомами образует нерегулярную соленоид-подобную структуру толщиной около 30 нанометров, так называемую 30 нм фибриллу. Дальнейшая упаковка этой фибриллы может иметь различную плотность. Если хроматин упакован плотно, его называют конденсированным или гетерохроматином, он хорошо видим под микроскопом. ДНК, находящаяся в гетерохроматине, не транскрибируется, обычно это состояние характерно для незначимых или молчащих участков. В интерфазе гетерохроматин обычно располагается по периферии ядра (пристеночный гетерохроматин). Полная конденсация хромосом происходит перед делением клетки. Если хроматин упакован неплотно, его называют эу- или интерхроматином. Этот вид хроматина гораздо менее плотный при наблюдении под микроскопом и обычно характеризуется наличием транскрипционной активности. Плотность упаковки хроматина во многом определяется модификациями гистонов — ацетилированием и фосфорилированием.
Считается, что в ядре существуют так называемые функциональные домены хроматина(ДНК одного домена содержит приблизительно 30 тысяч пар оснований), то есть каждый участок хромосомы имеет собственную «территорию». К сожалению, вопрос пространственного распределения хроматина в ядре изучен пока недостаточно. Известно, что теломерные (концевые) и центромерные (отвечающие за связывание сестринских хроматид в митозе) участки хромосом закреплены на белках ядерной ламины.
Ядерная оболочка, ядерная ламина и ядерные поры (кариолемма)
От цитоплазмы ядро отделено ядерной оболочкой, образованной за счёт расширения и слияния друг с другом цистерн эндоплазматической сети таким образом, что у ядра образовались двойные стенки за счёт окружающих его узких компартментов. Полость ядерной оболочки называется люменом или перинуклеарным пространством. Внутренняя поверхность ядерной оболочки подстилается ядерной ламиной, жёсткой белковой структурой, образованной белками-ламинами, к которой прикреплены нити хромосомной ДНК. Ламины прикрепляются к внутренней мембране ядерной оболочки при помощи заякоренных в ней трансмембранных белков — рецепторов ламинов. В некоторых местах внутренняя и внешняя мембраны ядерной оболочки сливаются и образуют так называемые ядерные поры, через которые происходит материальный обмен между ядром и цитоплазмой. Пора не является дыркой в ядре, а имеет сложную структуру, организованную несколькими десятками специализированных белков — нуклеопоринов. Под электронным микроскопом она видна как восемь связанных между собой белковых гранул с внешней и столько же с внутренней стороны ядерной оболочки.
![]()
Ядрышко находится внутри ядра, и не имеет собственной мембранной оболочки, однако хорошо различимо под световым и электронным микроскопом. Основной функцией ядрышка является синтез рибосом. В геноме клетки имеются специальные участки, так называемые ядрышковые организаторы, содержащие гены рибосомной РНК (рРНК), вокруг которых и формируются ядрышки. В ядрышке происходит синтез рРНК РНК полимеразой I, ее созревание, сборка рибосомных субчастиц. В ядрышке локализуются белки, принимающие участие в этих процессах. Некоторые из этих белков имеют специальную последовательность — сигнал ядрышковой локализации (NoLS, от англ. Nucleolus Localization Signal). Следует отметить, самая высокая концентрация белка в клетке наблюдается именно в ядрышке. В этих структурах было локализовано около 600 видов различных белков, причем считается, что лишь небольшая их часть действительно необходима для осуществления ядрышковых функций, а остальные попадают туда неспецифически.
Под электронным микроскопом в ядрышке выделяют несколько субкомпартментов. Так называемые Фибриллярные центры окружены участками плотного фибриллярного компонента, где и происходит синтез рРНК. Снаружи от плотного фибриллярного компонента расположен гранулярный компонент, представляющий собой скопление созревающих рибосомных субчастиц.
Ядерный матрикс
Ядерным матриксом некоторые исследователи называют нерастворимый внутриядерный каркас. Считается, что матрикс построен преимущественно из негистоновых белков, формирующих сложную разветвленную сеть, сообщающуюся с ядерной ламиной. Возможно, ядерный матрикс принимает участие в формировании функциональных доменов хроматина. В геноме клетки имеются специальные незначащие А-Т-богатые участки прикрепления к ядерному матриксу (англ. S/MAR — Matrix/Scaffold Attachment Regions), служащие, как предполагается, для заякоривания петель хроматина на белках ядерного матрикса. Впрочем, не все исследователи признают существование ядерного матрикса.

Эволюционное значение клеточного ядра
Основное функциональное отличие клеток эукариот от клеток прокариот заключается в пространственном разграничении процессов транскрипции (синтеза матричной РНК) и трансляции (синтеза белка рибосомой), что дает в распоряжение эукариотической клетки новые инструменты регуляции биосинтеза и контроля качества мРНК.
В то время, как у прокариот мРНК начинает транслироваться еще до завершения ее синтеза РНК-полимеразой, мРНК эукариот претерпевает значительные модификации (так называемый процессинг), после чего экспортируется через ядерные поры в цитоплазму, и только после этого может вступить в трансляцию. Процессинг мРНК включает несколько элементов.
Из предшественника мРНК (пре-мРНК) в ходе процесса, называемого сплайсингом вырезаются интроны — незначащие участки, а значащие участки — экзоны соединяются друг с другом. Причем экзоны одной и той же пре-мРНК могут быть соединены несколькими разными способами (альтернативный сплайсинг), так что один предшественник может превращаться в зрелые мРНК нескольких разных видов. Таким образом, один ген может кодировать сразу несколько белков.
Кроме того, интрон-экзонная структура генома, практически невозможная у прокариот (так как рибосомы смогут транслировать незрелые мРНК), дает эукариотам определенную эволюционную мобильность. Учитывая протяженность интронных участков, рекомбинация между двумя генами зачастую сводится к обмену экзонами. Благодаря тому, что экзоны часто соответствуют функциональным доменам белка, участки получившегося в результате рекомбинации «гибрида», зачастую сохраняют свои функции. В то же время у прокариот рекомбинация между генами невозможна без разрыва в значащей части, что безусловно уменьшает шансы на то, что получившийся белок будет функционален.
Модификациям подвергаются концы молекулы мРНК. К 5′ -концу молекулы прикрепляется 7-метилгуанин (так называемый кэп). К 3′-концу нематрично присоединяются несколько десятков остатков аденина (полиаденирование).
Процессинг мРНК тесно сопряжен с синтезом этих молекул и необходим для контроля качества. Непроцессированная или не полностью процессированная мРНК не сможет выйти из ядра в цитоплазму или будет нестабильна и быстро деградирует. У прокариот нет таких механизмов контроля качества, и из-за этого прокариотические мРНК имеют меньший срок жизни — нельзя допустить, чтобы неправильно синтезированная молекула мРНК, если такая появится, транслировалась в течение долгого времени.
Происхождение ядра
Клеточное ядро является важнейшей чертой эукариотических организмов, отличающей их от прокариот и архей. Несмотря на значительный прогресс в цитологии и молекулярной биологии, происхождение ядра не выяснено и является предметом научных споров. Выдвинуто 4 основных гипотезы происхождения клеточного ядра, но ни одна из них не получила широкой поддержки. [1]
Гипотеза, известная как «синтропная модель», предполагает что ядро возникло в результате симбиотических взаимоотношений между археей и бактерией (ни археи, ни бактерии не имеют оформленных клеточных ядер). По этой гипотезе, симбиоз возник, когда древняя архея (сходная с современными метаногенными археями), проникла в бактерию (сходную с современными Миксобактериями). Впоследствии архея редуцировалась до клеточного ядра современных эукариот. Эта гипотеза аналогична практически доказанным теориям происхождения митохондрий и хлоропластов, которые возникли в результате эндосимбиоза прото-эукариот и аэробных бактерий. [2] Доказательством гипотезы является наличие одинаковых генов у эукариот и архей, в частности генов гистонов. Также миксобактерии быстро передвигаются, могут образовывать многоклеточные структуры и имеют киназы и G-белки, близкие к эукариотическим. [3]
Согласно второй гипотезе, прото-эукариотическая клетка эволюционировала из бактерии без стадии эндосимбиоза. Доказательством модели является существование современных бактерий из отряда Planctomycetes, которые имеют ядерные структуры с примитивными порами и другие клеточные компартменты, ограниченные мембранами (ничего похожего у других прокариот не обнаружено). [4]
Согласно гипотезе вирусного эукариогенеза, окруженное мембраной ядро, как и другие эукариотические элементы, произошли вследствие инфекции прокариотической клетки вирусом. Это предположение основывается на наличии общих черт у эукариот и некоторых вирусов, а именно геноме из линейных цепей ДНК, кэпировании мРНК и тесном связывании генома с белками (гистоны эукариот принимаются аналогами вирусных ДНК-связывающих белков). По одной версии, ядро возникло при фагоцитировании (поглощении) клеткой большого ДНК-содержащего вируса. [5] По другой версии, эукариоты произошли от древних архей, инфицированных поксвирусами. Это гипотеза основана на сходстве ДНК-полимеразы современных поксвирусов и эукариот. [6] [7] Также предполагается, что нерешенный вопрос о происхождении пола и полового размножения может быть связан с вирусным эукариогенезом. [8]
Наиболее новая гипотеза, названная экзомембранной гипотезой, утверждает, что ядро произошло от одиночной клетки, которая в процессе эволюции выработала вторую внешнюю клеточную мембрану; первичная клеточная мембрана после этого превратилась в ядерную мембрану, и в ней образовалась сложная система поровых структур (ядерных пор) для транспорта клеточных компонентов, синтезированных внутри ядра. [9]
dic.academic.ru
Имеют ли бактерии ядро, или Особенности строения прокариотических клеток
Тот факт, что бактерии вместе с археями были отнесены биологами к прокариотам, позволяет сделать некоторые выводы об особенностях строения этих микроорганизмов. В частности, имеется возможность ответить на вопрос о том, имеют ли бактерии такое же ядро, как и многие другие живые организмы.
Основное их отличие от эукариот – это то, что бактерии не имеют ядра. Бактериальным клеткам вообще не свойственны развитые внутриклеточные мембранные структуры. В клетке цианобактерии можно обнаружить небольшие мембранные образования, напоминающие пузырьки и названные тилакоидами. В них собраны системы, осуществляющие фотосинтез, – пигменты и ферментный комплекс. Эти микроорганизмы, признанные наиболее эволюционно продвинутыми, осуществляют процесс фотосинтеза аналогично эукариотам – организмам, клетки которых имеют настоящее, оформленное ядро.
Небольшие мембранные образования помогают бактериальным клеткам организовывать основные процессы, обеспечивающие их существование.
Если сравнивать их по функциям с органеллами эукариотических клеток, то можно найти примитивный аппарат Гольджи, митохондрии, ЭПС (эндоплазматическую сеть). Однако бактерии не имеют настоящего ядра, окруженного мембраной. Все бактерии имеют нуклеоид, а не ядро – кольцевую молекулу ДНК, свободно расположенную в цитоплазме.
Бактериальная клетка: особенности
Форму бактерии определяет клеточная стенка. Ее размер вместе с капсулой в некоторых случаях может быть больше, чем расположившаяся внутри клетка. Стенка имеет избирательную проницаемость и способна пропускать внутрь необходимые вещества и выводить из нее продукты метаболизма. Снаружи нее часто можно обнаружить жгутики или ворсинки – выпячивания мембраны, позволяющие организму самопроизвольно передвигаться.
Наличие клеточной стенки характерно для группы бактерий, которые называют грамположительными. Под клеточной стенкой расположена мембрана. А вокруг молекулы ДНК она отсутствует, и это позволяет утверждать, что бактерии не имеют оформленного мембраной ядра.
Цитоплазма
Под этой сложной оболочкой бактерии находится цитоплазма – гелевая масса различной плотности, в толще которой находятся включения:
- рибосомы, продуцирующие белок;
- небольшие мембранные структуры;
- жировые включения (гликоген);
- полифосфатные соединения (волютин);
- полисахариды;
- бета-оксимасляная кислота.
Состав включения зависит от потребности бактерии в источниках энергии и питательных веществах. Некоторые бактерии имеют цитоскелет – систему трубочек, способную ориентировать внутри клетки ее основные компоненты. В частности, они позволяют правильно располагаться молекуле ДНК во время репликации, несмотря на то, что бактерии не имеют в клетке настоящего ядра и гистонов.
Примерно по центру клетки обнаруживается нуклеоид – место расположения наследственной информации. Оформленного ядра, которое бы имело собственную мембрану, основные белки (гистоны) и ферментный комплекс, принимающий участие в воспроизведении наследственной информации и ее реализации, у бактерии нет.
Отсутствие оформленного ядра определяет простой процесс воспроизведения генетической информации – кольцевая молекула ДНК просто удваивается перед делением клетки, и по одной копии оказывается в дочерних организмах.
Однако существует особенность передачи генетической информации, которая делает бактерии уникальными для генетиков и молекулярных биологов. Возможность их функционирования как раз связана с тем, что бактерии не имеют в клетке ядра. Внутри клеток обнаружены нехромосомные элементы, способные к передаче информации в обход ядра. Наиболее изучены среди них такие:
- Плазмиды.
- Транспозоны и IS-элементы (вставные последовательности).
- Умеренные фаги.
Любопытно, что размер генетической информации, обнаруженный в мобильных элементах, существенно превышает ее число в главной молекуле ДНК. Именно они имеют прямое отношение к:
- защитным реакциям бактерий,
- их быстрому привыканию к лекарственных препаратам,
- способности синтезировать антибиотики и необычные для бактерий сахара и использовать для питания некоторые не свойственные для своего вида источники.
Ничего подобного плазмидам бактерий у эукариотических организмов нет, поскольку они имеют оформленное ядро, препятствующее контакту основного генома с неядерными элементами. Они способны к самостоятельному воспроизведению и имеют для этого собственный набор необходимых генов.
Высокая изменчивость была причиной того, что биологи длительное время считали, что у них нет такого понятия, как вид. Только появление чистых культур позволило сделать вывод о том, что это понятие вполне применимо к этим организмам, и местом локализации основного генома у них является их примитивное ядро или нуклеоид.
Таким образом, бактерии не имеют ядра, и это позволяет им обмениваться генетической информации «по горизонтали», быстро перенося полезные гены внутри существующей популяции клеток и существенно повышая их адаптивность к изменениям окружающей среды.
Клетки архей – варианты безъядерного существования
probakterii.ru