
Оглавление:
Закономерности моногенного и полигенного наследования
Страницы работы


Содержание работы
ТЕМА: «ЗАКОНОМЕРНОСТИ МОНО- И ПОЛИГЕННОГО НАСЛЕДОВАНИЯ»
I. Научно-методическое обоснование
Полученные знания и умения необходимы студентам при изучении патологической физиологии, психиатрии, нервных, детских и внутренних болезней, при работе врачом общего профиля и медико-генетической консультации.
1. Научить студентов решать задачи на моно-, ди- и полигибридное скрещивание, анализирующее скрещивание, полигенное наследование.
2. Проверить степень усвоения студентами знаний по закономерностям моно- и полигенного наследования признаков.
1. Знать типы и виды наследования, сущность законов Менделя при моно- и полигибридном скрещивании, причинно-следственные связи в закономерностях наследования признаков.
2. Уметь решать задачи на взаимодействие аллельных и неаллельных генов.
3. Ознакомиться со статистическим характером закономерностей наследования.
IY. Содержание, хронокарта занятия
п/п│ Этапы занятия │в мин.
1. Вступительное слово. Ответы на вопросы. 5
2. Проверка знаний студентов по контрольным вопросам темы
методом фронтального опроса. 50
2.1 Наследование. Типы и виды при моногенном и полигенном
2.2 Гибридологический анализ, его сущность.
2.3 Моногенное наследование признаков, его закономерности:
— закономерности наследования при моногибридном скре-
щивании (закон единообразия гибридов первого поколе-
ния, закон расщепления, закон «чистоты гамет» У.Бэтсо-
на и его цитологические основы; анализирующее скрещи-
вание прямое и возвратное);
— закономерности наследования при ди- и полигибридном
скрещивании (закон независимого комбинирования неал-
лельных генов и его цитологические основы); решетка
Пеннета; фенотипический радикал;
— условия проявления законов Г.Менделя, их статисти-
— менделирующие признаки, условия их проявления; мен-
делирующие признаки человека.
2.4 Полигенное наследование признаков, условия его проявле-
3. Тестовый контроль. 10
4. Подведение итогов опроса. Объяснение ООД по выполнению
лабораторной работы. 5
5. Выполнение лабораторной работы. 50
6. Проверка выполнения лабораторной работы и оценка знаний
7. Подведение итогов занятия. Объяснение ООД по очередной
Y. Методика проведения и достижения учебно-целевых задач
Провести учет посещаемости, обратить внимание на воспитательные моменты. Указать цель занятия, его мотивационную характеристику. Ответить на вопросы студентов по теме занятия. Провести коллективный разбор вопросов занятия с участием студентов всей группы.
При разборе вопроса 2.3 законы единообразия и расщепления признаков при моногенном наследовании можно разобрать по таблице 1-4.
По вопросу 2.4 полигенное наследование разобрать по таблицам 6-7.
При выполнении лабораторной работы студенты должны решить задачи из задачника Э. А. Каминской на моногибридное скрещивание NN 11, 12, 13, 16, 17, 53, 54; на дигибридное скрещивание — NN 119, 120, 121, 122, 123, 131; на полигенное наследование — N 165.
Подвести итог занятия. Особое внимание обратить на правильность решения и оформления ситуационных задач на закономерности моно- и полигенного наследования.
Дать ООД к очередному занятию.
YI. Методическое обеспечение занятия
1. Классификация типов наследования (I) (31).
2. Классификация типов наследования (II) (32).
3. Аутосомное наследование (неполное доминирование) (34).
4. Аутосомное наследование (полное доминирование) (35).
5. Аутосомное наследование (кодоминирование) (36).
6. Полимерия (I): наследование признака при неаллельных генах (38).
7. Полимерия (II): наследование признака при неаллельных генах (38).
1. (256) Моногибридное скрещивание.
2. (257) Гибридологический анализ при взаимодействии генов.
3. (263) Закон независимого комбинирования признаков (III з-н Менделя / Поведение гомологичных хромосом при дигибридном скрещивании.
4. (268) Определение гетерозиготы.
5. (270) Дигибридное скрещивание.
6. (274) Анализирующее скрещивание.
7. (275) Термины и условные обозначения, принятые в генетике.
8. (277) Г. Мендель, Э. Чермах, К. Корренс, Г.де Фриз.
9. (278) Поведение гомологичных хромосом в анафазу I.
10. (279) Характер расщепления у кукурузы в F 42 0 / Наследование признака окраски раковин у улиток.
11. (285) Составление решетки Пеннета / Доминирование.
12. (287) Результаты гибридизации двух разновидностей ячменя.
13. (288) Доминантные и рецессивные признаки. Явление доминирования.
14. (289) Неполное доминирование. 290. Закон расщепления Менделя.
15. (291) Определение фенотипа, гомозиготы, гетерозиготы. Закономерности моногибридного скрещивания.
16. (295) Вопросы по скрещиванию / Мичуринская теория доминирования.
17. (296) Закон независимого наследования признаков.
18. (297) Полимерия.
19. (332) Промежуточный характер признака. Схема расщепления.
20. (336) Отдаленная гибридизация.
21. (338) Определение генетики. Памятник Менделю в г. Брно.
22. (339) Схема гибридизации.
23. (345) Тригибридное и полигибридное скрещивание.
24. (347) Правило единообразия первого поколения гибридов.
25. (348) Гипотеза частоты гамет.
26. (290) Закон расщепления Менделя.
1. Диапроектор Пеленг-800.
2. Биология. Под ред. Ярыгина В. Н., М.: Медицина, 1984, стр.81-83, 94-103.
3. Слюсарев А. А., Жукова С. В. Биология. Киев: Вища школа, 1987, стр.68-71, 73-75.
4. Основы общей биологии. Под ред. Либберта Э. М., Мир, с.201-202, 305-308.
5. Каминская Э. П. Сборник задач по генетике. Минск, Вышэйшая школа, 1977, с.5-11, 14-26.
1. Гершензон С. М. Основы современной генетики. Киев, Навукова думка, 1979, с.24-59.
2. Бочков Н. И., Захаров А. Ф., Иванов В. И. Медицинская генетика. М., Медицина, 1984, с.55-61.
3. Хелевин Н. В. с соавт. Задачник по общей и медицинской генетике. М., Высшая школа, 1984, с.13-16, 22-26, 61.
4. Бердышев Г. Д., Криворучко И. Ф. Генетика человека с основами медицинской генетики. Киев, Вища школа, 1979, с.207-208.
vunivere.ru
Справочник химика 21
Химия и химическая технология
Аддитивное полигенное наследование


В предшествующем разделе генетический анализ количественного признака на биометрическом уровне обсуждался в отношении нормальных признаков с унимодальным и почти нормальным распределением в популяции. Было показано, что простая модель аддитивною полигенного наследования удовлетворяет этим свойствам, и тем самым корреляции родитель-ребенок и сибс-сибс можно использовать для оценки наследуемости. [c.249]
У человека известно несколько заболеваний, например, склонность к гастритам, в основе которых лежит полигенное наследование, то есть аддитивное действие целого ряда генов. Для таких заболеваний характерно, что частота их у близких родственников больного (пробанда) значительно выше, а при снижении степени родства она быстро падает. Эти заболевания возникают у [c.77]
С другой стороны, как показано в разд. 3.6.1.3, эти свойства не исключают наличия эффекта главного гена с простым типом наследования, который действует на фоне аддитивно-полигенной системы. Биологически вполне правдоподобно, что лишь несколько главных генов могут быть основными генетическими факторами ряда заболеваний, проявляющими свои эффекты на фоне многих генов, менее значимых для патогенеза этих заболеваний. [c.239]
Сложный функциональный дефект вызывается комбинацией малых нарушений. Как упоминалось выше, аддитивно-полигенная модель, используемая для анализа мультифакториального наследования, является слишком упрощенной абстракцией. В действительности изменчивость не одномерна, и к определенному заболеванию может [c.258]
Все распределения на рис. 3.55 имеют только одну моду (т. е. они унимодальны). Кроме того, они сходны с нормальным распределением. Это сходство увеличивается с возрастанием числа рассматриваемых генов (и), т. е. при возрастании п нормальное распределение является предельным случаем биномиального. Можно показать, что эта аппроксимация становится удовлетворительной как раз тогда, когда частоты положительных и отрицательных аллелей не равны. Чем ближе к симметрии, тем большие значения п требуются для достижения той же степени аппроксимации. Вообще, унимодальное распределение, которое более или менее точно аппроксимируется нормальным, является типичным для генетической модели аддитивной полигении. Однако ни унимодальность распределения, ни его форма не зависят от конкретных свойств этой модели (равных и аддитивных вкладов генов) и потому могут служить индикаторами мультифакториального наследования в более общем смысле. [c.239]
Результаты семейных исследований этих заболеваний не согласуются с простым менделевским наследованием. Анализ, основанный на различных полигенных моделях, позволил сделать вывод о том, что в этиологию этих болезней вовлечены многие неспецифические гены, действующие вместе со средовыми факторами. Первичный биологический эффект этих генов остается неизвестным и рассматривается как черный ящик . Обычно считают, что количество генов относительно велико и что вклад в патогенез болезни каждого из вовлеченных индивидуальных генов относительно мал, т.е. предполагается аддитивное действие этих генов. В случаях когда болезнь проявляется как качественный признак с двумя альтернативными классами норма и пораженные (например, как при врожденных уродствах), предполагается наличие порога. Считается, что если сумма факторов, действующих на индивид, превышает этот порог, то заболевание проявляется. В других случаях, когда число генов недостаточно и значение подверженности индивида оказывается меньше порогового, но вблизи него, это может проявляться не как болезнь, а скорее как отклонение. [c.298]
Известно, что наследование некоторых признаков характеризуется такими генетическими феноменами, как гетерогенность, неполная пенетрантность или интегрирующее взаихмодействие. Наиболее сложное наследование — полигенное, при котором аллели нескольких разных локусов взаимодействуют друг с другом по принципу аддитивности, обусловливая таким образом риок проявления признака или заболевания. Каждый добавочный плохой аллель увеличивает предрасположенность индивида, но ни один из локусов не является существенным для [c.233]
Генетика человека Т.3 (1990) — [ c.241 , c.249 ]
chem21.info
Полигенный тип наследования. Формы взаимодействия неаллельных генов
Типы наследования признаков. Моногенный тип наследования. Формы взаимодействия аллельных генов.
Цитоплазматическая наследственность. Плазмиды, группы плазмид.
Цитоплазматическая наследственность. Она обусловлена ДНК органелл цитоплазмы (митохондрии, пластиды, клеточный центр). Внехромосомные генетические элементы перечисленных органелл способны автономно существовать и удваиваться.
Плазмиды – внехромосомные генетические элементы, способные существовать и размножаться в клетке автономно от геномной ДНК.
Моногенный тип наследования – доминантный признак контролируется парой или несколькими парами аллельных генов (1 пара аллельных генов контролирует 1 признак).
Он делится на:
Полигенное наследование – наследование при кот несколько генов определяют проявление одного признака.
Плейотропия – наследование при кот 1 пара генов отвечает за развитие нескольких признаков
Аллельные гены — это гены, расположенные в одинаковых участках или локусах двух гомологичных хромосом.
Формы взаимодействия аллельных генов:
— Полное доминирование – один аллельный ген полностью подавляет действие другого аллельного гена.
— Неполное доминирование – доминантный ген не полностью подавляет рецессивный.
— Сверхдоминирование – когда в гетерозиготном организме рецессивный ген усиливает действие доминантного гена, при этом наблюдается явление гетерозиса или гибридной силы.
— Кодоминирование – два аллельных гена одновременно проявляют своё действие в фенотипе.
Полигенный тип наследования – это такой тип наследования, который контролируется несколькими парами неаллельных генов, т.е. один признак – несколько генов.
Неаллельные гены –это гены, которые находятся в разных парах хромосом или в разных локусах гомологичных хромосом и отвечают за развитие одного или нескольких признаков.
Формы взаимодействия неаллельных генов:
1) эпистаз – подавление дейтсвия или проявления генов генами другого аллеля. Различают эпистаз доминантный (доминантный ген одного аллеля подавляет действие другого доминантного неаллельного ему гена. Ген подавляющий – эпистатический, подавляемый – гипостатический. При дигибридном скрещивании F2 12:3:1 или F2 13:3 и т.д.) и рецессивный (рецессивный ген в гомозиготном состоянии подавляет действие доминантного гена другой аллели. Расщепление в F2 9:3:4)
— Комплементарность – это когда два неаллельных, доминантных гена, сшедшиеся в одном генотипе, дают новое проявление признака, которое несвойственно для каждого гена в отдельности.
— Полимерия – один признак контролируется несколькими парами неаллельных генов.
studopedia.ru
НАСЛЕДОВАНИЕ ПОЛИГЕННОЕ
Термины и определения, используемые в селекции, генетике и воспроизводстве сельскохозяйственных животных. — М.: ВНИИплем . И. М. Дунин . 1996 .
Смотреть что такое «НАСЛЕДОВАНИЕ ПОЛИГЕННОЕ» в других словарях:
наследование полигенное — (греч. poly много + гены) Н. признака, контролируемого совместно группой неаллельных генов … Большой медицинский словарь
Наследование полигенное полигения — Наследование полигенное, полигения * наследаванне палігеннае, палігенія * polygenic inheritance or polygeny наследование признаков, которые определяются многими генами, обладающими в отдельности слабым действием. Фенотипическое проявление… … Генетика. Энциклопедический словарь
полигенное наследование — Наследование количественных признаков, т.е. признаков, выражение которых определяется взаимодействием значительного числа генов (полигенов). [Арефьев В.А., Лисовенко Л.А. Англо русский толковый словарь генетических терминов 1995 407с.] Тематики… … Справочник технического переводчика
наследование — Передача генетической информации одним поколением другому (родителями или родителем потомству); типы и характер Н. зависят от характера воспроизведения генетического материала (удвоение и распределение), от локализации генов (ядерная,… … Справочник технического переводчика
полигенное наследование — polygenic inheritance полигенное наследование. Hаследование количественных признаков, т.е. признаков, выражение которых определяется взаимодействием значительного числа генов (полигенов). (Источник: «Англо русский толковый словарь генетических… … Молекулярная биология и генетика. Толковый словарь.
полигенное наследование — poligeninis paveldėjimas statusas T sritis augalininkystė apibrėžtis Paveldėjimas požymio, kurį nulemia daugelis kartu veikiančių genų. atitikmenys: angl. polygeny; polygenic inheritance rus. полигения; полигенное наследование ryšiai: sinonimas – … Žemės ūkio augalų selekcijos ir sėklininkystės terminų žodynas
наследование — inheritance наследование. Передaчa генетической информации одним поколением другому (родителями или родителем потомству); типы и характер Н. зависят от характера воспроизведения генетического материала (удвоение и распределение), от локализации… … Молекулярная биология и генетика. Толковый словарь.
СКЛЕРОЗ РАССЕЯННЫЙ — мед. Рассеянный склероз (PC) хроническое демиелинизирующее заболевание головного и спинного мозга, характеризующееся развитием рассеянных (во времени и пространстве) очагов демиелинизации и множественных неврологических симптомов; заболевание… … Справочник по болезням
Диате́зы — (diatheses: греч. diathesis предрасположение, склонность к чему либо) аномалии конституции, характеризующиеся предрасположенностью к некоторым болезням и определенному типу неадекватных реакций на обычные раздражители. Учение о Д. окончательно не … Медицинская энциклопедия
Наследственность — I Наследственность присущее всем организмам свойство повторять в ряду поколений одинаковые признаки и особенности развития; обусловлено передачей в процессе размножения от одного поколения к другому материальных структур клетки,… … Большая советская энциклопедия
agricultural_terms.academic.ru
Моногенное и полигенное наследование. Особенности аутосомного и сцепленного с полом наследования
Признак становится реальностью тогда, когда в цитоплазме появляется продукт активности данного гена. Простой признак развивается как результат определенного типа взаимодействия соответствующих аллельных генов (моногенное наследование). Большинство признаков контролируется совместным действием многих генов (полигенное наследование).

Аутосомное наследование. Характерные черты аутосомного наследования признаков обусловлены тем, что соответствующие гены, расположенные в аутосомах, представлены у всех особей вида в двойном наборе. Это означает, что любой организм получает такие гены от обоих родителей. В соответствиис законом чистоты гамет в ходе гаметогенеза все половые клетки получают по одному гену из каждой аллельной пары (рис. 6.6). Обоснованием этого закона является расхождение гомологичных хромосом, в которых располагаются аллельные гены, к разным полюсам клетки в анафазе I мейоза.
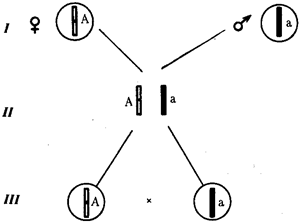
Рис. 6.6. Обоснование закономерностей аутосомного наследования признаков:
I — гаплоидные гаметы родителей, II — диплоидный генотип особи (фенотип зависит от взаимодействия аллельных генов Аа); III— гаплоидные гаметы гетерозиготной особи (гаметы «чисты», так как несут по одному из пары аллельных генов); черным и белым обозначены гомологичные хромосомы; буквами — определенные локусы
Ввиду того что развитие признака у особи зависит в первую очередь от взаимодействия аллельных генов, разные его варианты, определяемые разными аллелями соответствующего гена, могут наследоваться по аутосомно-доминантному или аутосомно-рецессивному типу, если имеет место доминирование. Возможен также промежуточный тип наследования признаков при других видах взаимодействия аллелей (см. разд. 3.6.5.2).
При доминировании признака, описанном Г. Менделем в его опытах на горохе, потомки от скрещивания двух гомозиготных родителей, различающихся по доминантному и рецессивному вариантам данного признака, одинаковы и похожи на одного изних (закон единообразия F1). Описанное Менделем расщепление по фенотипу в F2 в отношении 3:1 в действительности имеет место лишь при полном доминировании одного аллеля над другим, когда гетерозиготы фенотипически сходны с доминантными гомозиготами(закон расщепления в F2).

Рис. 6.7. Аутосомное наследование признака:
I — полное доминирование (наследование цвета лепестков у гороха); II — неполное доминирование (наследование цвета лепестков у ночной красавицы)
Наследование рецессивного варианта признака характеризуется тем, что он не проявляется у гибридов F1, а в F2 проявляется у четверти потомков (рис. 6.7.7).
В случаях формирования у гетерозигот нового варианта признака по сравнению с гомозиготами, что наблюдается при таких видах взаимодействия аллельных генов, как неполное доминирование, кодоминирование, межаллельная комплементация, гибриды F1 не похожи на родителей, а в F2 образуется три фенотипических группы потомков (рис. 6.7, II).
Завершая описание характерных черт аутосомно-доминантного и аутосомно-рецессивного наследования, уместно напомнить, что хотя в случае доминирования одного из аллелей присутствие в генотипе другого, рецессивного, аллеля не сказывается на формировании доминантного варианта признака, на фенотипическое проявление аллеля оказывает влияние вся система генотипа конкретного организма, а также среда, в которой реализуется наследственная информация. В связи с этим существует возможность неполной пенетрантности доминантного аллеля у особей, имеющих его в генотипе.

Рис. 6.8. Наследование признака окраски глаз у дрозофилы:
I, II — различие результатов скрещивания в зависимости от пола родителя с доминантным признаком; зачернены отцовские хромосомы
Сцепленное с полом наследование. Анализ наследования признака окраски глаз у дрозофилы в лаборатории Т. Моргана выявил некоторые особенности, заставившие выделить в качестве отдельного типа наследования признаков сцепленное с полом наследование (рис. 6.8).
Зависимость результатов эксперимента от того, кто из родителей являлся носителем доминантного варианта признака, позволила высказать предположение, что ген, определяющий окраску глаз у дрозофилы, расположен в Х-хромосоме и не имеет гомолога в Y-хромосоме. Все особенности сцепленного с полом наследования объясняются неодинаковой дозой соответствующих генов у представителей разного — гомо- и гетерогаметного пола.
Гомогаметный пол несет двойную дозу генов, расположенных в Х-хромосоме. Развитие соответствующих признаков у гетерозигот (Х A Х a ) зависит от характера взаимодействия между аллельными генами. Гетерогаметный пол имеет одну Х-хромосому (ХО или XY). У некоторых видов Y-хромосома генетически инертна, у других она содержит некоторое количество структурных генов, часть из которых гомологична генам Х-хромосомы (рис. 6.9). Гены негомологичных участков Х- и Y-хромосом (или единственной Х-хромосомы) у гетерогаметного пола находятся в гемизиготном состоянии. Они представлены единственной дозой: Х A Y, Х a Х, XY B . Формирование таких признаков у гетерогаметного пола определяется тем, какой аллель данного гена присутствует в генотипе организма.
Характер наследования сцепленных с полом признаков в ряду поколений зависит от того, в какой хромосоме находится соответствующий ген. В связи с этим различают Х-сцепленное и Y-сцепленное (голандрическое) наследование.
Х-сцепленное наследование. Х-хромосома присутствует в кариотипе каждой особи, поэтому признаки, определяемые генами этой хромосомы, формируются у представителей как женского, так и мужского пола. Особи гомогаметного пола получают эти гены от обоих родителей и через свои гаметы передают их всем потомкам. Представители гетерогаметного пола получают единственную Х-хромосому от гомогаметного родителя и передают ее своему гомогаметному потомству.
У млекопитающих (в том числе и человека) мужской пол получает Х-сцепленные гены от матери и передает их дочерям. При этом мужской пол никогда не наследует отцовского Х-сцепленного признака и не передает его своим сыновьям (рис. 6.10).
Так как у гомогаметного пола признак развивается в результате взаимодействия аллельных генов, различают Х-сцепленное доминантное и Х-сцепленное рецессивное наследование. Х-сцепленный доминантный признак (красный цвет глаз у дрозофилы) передается самкой всему потомству. Самец передает свой Х-сцепленный доминантный признак лишь самкам следующего поколения. Самки могут наследовать такой признак от обоих родителей, а самцы — только от матери.

Рис. 6.9. Схема гомологичных и негомологичных локусов
в половых хромосомах человека:
I — Х-хромосомы: заштрихованы локусы, отсутствующие в Y-хромосоме (красно-зеленая слепота, гемофилия и др.); II — Y-хромосома: заштрихованы локусы, отсутствующие в Х-хромосоме (перепонки между пальцами, гены-детерминаторы развития организма по мужскому типу); участки Х- и Y-хромосом, соответствующие гомологичным локусам, не заштрихованы
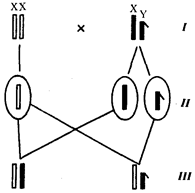
Рис. 6.10. Обоснование закономерностей сцепленного с полом
I — сочетание половых хромосом в кариотипе представителей разного пола; II — гомогаметный пол образует один тип гамет, гетерогаметный — два; III — представители гомогаметного пола получают хромосомы от обоих родителей; представители гетерогаметного пола получают Х-хромосому от гомогаметного родителя, а Y-хромосому — от гетерогаметного родителя; это справедливо для генов, расположенных в негомологичных локусах Х- и Y-хромосом; зачернены отцовские хромосомы
Х-сцепленный рецессивный признак, (белый цвет глаз у дрозофилы) у самок проявляется только при получении ими соответствующего аллеля от обоих родителей (X a X a ). У самцовX a Y он развивается при получении рецессивного аллеля от матери. Рецессивные самки передают рецессивный аллель потомкам любого пола, а рецессивные самцы —только «дочерям» (см. рис. 6.8).
При Х-сцепленном наследовании, так же как и при аутосомном, возможен промежуточный характер проявления признака у гетерозигот. Например, у кошек пигментация шерсти контролируется Х-сцепленным геном, разные аллели которого определяют черную (X A и рыжую (X A’ ) пигментацию. Гетерозиготные самки X A X A’ имеют пеструю окраску шерсти. Самцы же могут быть либо черными (X A Y, либо рыжими (X A’ Y).
Голандрическое наследование. Активно функционирующие гены Y-хромосомы, не имеющие аллелей в Х-хромосоме, присутствуют в генотипе только гетерогаметного пола, причем в гемизиготном состоянии. Поэтому они проявляются фенотипически и передаются из поколения в поколение лишь у представителей гетерогаметного пола. Так, у человека признак гипертрихоза ушной раковины («волосатые уши») наблюдается исключительно у мужчин и наследуется от отца к сыну.
Дата добавления: 2014-11-12 ; просмотров: 1613 . Нарушение авторских прав
studopedia.info